![]()
In a new review article published today in Journal of Venomous Animals and Toxins including Tropical Diseases (JVATiTD), Anna Nekaris and colleagues review the remarkable biochemistry and etiology of slow loris venom.

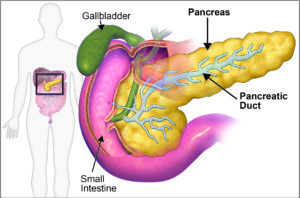 Figure 1: The pancreas is part of the digestive system, note the connection of the pancreas to the intestine through the pancreatic duct. Image adapted from Blausen.com staff (2014).[/caption]
Malfunction of these cells is associated with a series of pancreatic diseases. Pancreatic cancer can affect any of these cell types and has the lowest survival rate in the UK among all cancer types. Diabetes is the most common disease of the endocrine compartment and affects over 4 million people in the UK. Other serious diseases, although perhaps less well known, include pancreatitis (inflammation of the pancreas), cystic fibrosis (which affects ductal cells) and pancreatic insufficiency (reduced acinar cell function). Severe cases of pancreatic disease often leave patients with the need for a pancreas or islet cell transplant deriving from deceased organ donors. However, the greatest hurdles to pancreas transplants include a limited number of organ donors and a need for life-time immunosuppression.
We aimed to produce a 3D culture system for long-term expansion of hPOs to serve as the basis for studies of pancreas ductal cells, pancreatic diseases and the development of cell therapy for diabetes. Whilst developing the hPO model we were aware that for a cell therapy to translate into the clinic it must meet certain criteria which include cell expansion in a chemically defined manner that is compatible with Good Manufacturing Processes (GMP) with strong evidence that the cells pose no risk of tumour formation.
To generate hPOs, we isolated ductal fragments from small samples of human pancreatic tissue and placed these in an ECM to act as a 3D scaffold. In order for this to be successful we had to develop a culture medium which would include all the signals and growth factors required to support the transition of pancreatic ducts into expandable pancreas organoids (Figure 2). Through a series of optimisations and iterations in which we tested activators and inhibitors of key cell signalling pathways we achieved a chemically defined culture medium that sustains the long-term culture of the pancreas cells. In addition, we developed a protocol to freeze the pancreatic tissue and recover healthy long-term expanding organoids from it. This gives us the ability to bank tissue, share samples and derive organoids at a later date.
[caption id="attachment_11730" align="aligncenter" width="654"]
Figure 1: The pancreas is part of the digestive system, note the connection of the pancreas to the intestine through the pancreatic duct. Image adapted from Blausen.com staff (2014).[/caption]
Malfunction of these cells is associated with a series of pancreatic diseases. Pancreatic cancer can affect any of these cell types and has the lowest survival rate in the UK among all cancer types. Diabetes is the most common disease of the endocrine compartment and affects over 4 million people in the UK. Other serious diseases, although perhaps less well known, include pancreatitis (inflammation of the pancreas), cystic fibrosis (which affects ductal cells) and pancreatic insufficiency (reduced acinar cell function). Severe cases of pancreatic disease often leave patients with the need for a pancreas or islet cell transplant deriving from deceased organ donors. However, the greatest hurdles to pancreas transplants include a limited number of organ donors and a need for life-time immunosuppression.
We aimed to produce a 3D culture system for long-term expansion of hPOs to serve as the basis for studies of pancreas ductal cells, pancreatic diseases and the development of cell therapy for diabetes. Whilst developing the hPO model we were aware that for a cell therapy to translate into the clinic it must meet certain criteria which include cell expansion in a chemically defined manner that is compatible with Good Manufacturing Processes (GMP) with strong evidence that the cells pose no risk of tumour formation.
To generate hPOs, we isolated ductal fragments from small samples of human pancreatic tissue and placed these in an ECM to act as a 3D scaffold. In order for this to be successful we had to develop a culture medium which would include all the signals and growth factors required to support the transition of pancreatic ducts into expandable pancreas organoids (Figure 2). Through a series of optimisations and iterations in which we tested activators and inhibitors of key cell signalling pathways we achieved a chemically defined culture medium that sustains the long-term culture of the pancreas cells. In addition, we developed a protocol to freeze the pancreatic tissue and recover healthy long-term expanding organoids from it. This gives us the ability to bank tissue, share samples and derive organoids at a later date.
[caption id="attachment_11730" align="aligncenter" width="654"]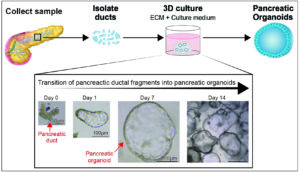 Figure 2: Human pancreatic ducts can be cultured in vitro and expanded as pancreatic organoids.[/caption]
Importantly, genetic analysis showed that even after long-term expansion the hPOs did not show large genetic abnormalities. This is crucial, as accumulation of genetic defects during culturing, can alter cellular behaviours and impact on the results. We also found that upon transplant of these cells into mouse pancreas the hPO cells do not form tumours, indicating their genetic safety both in vitro and in vivo.
Finally, we aimed to develop a chemically defined ECM in which to culture hPOs as the commonly used ECMs (Matrigel and BME 2-RGF) are animal-derived and are unsuitable for cell therapies. In collaboration with Cellendes, a company specialised in hydrogel technology, we could show for the first time the derivation and expansion of hPOs in a fully chemically defined hydrogel.
Our demonstration of the long-term expansion of human pancreas organoids in chemically defined conditions makes this an exciting system for many areas of research (Figure 3). This system can be used as a model of healthy ductal pancreas biology and the development of disease models of the ductal compartment, such as cystic fibrosis. Organoids can also be derived from diseased states and have already been used to model different types of pancreatic cancer (PDAC and IPMN).
[caption id="attachment_11731" align="aligncenter" width="605"]
Figure 2: Human pancreatic ducts can be cultured in vitro and expanded as pancreatic organoids.[/caption]
Importantly, genetic analysis showed that even after long-term expansion the hPOs did not show large genetic abnormalities. This is crucial, as accumulation of genetic defects during culturing, can alter cellular behaviours and impact on the results. We also found that upon transplant of these cells into mouse pancreas the hPO cells do not form tumours, indicating their genetic safety both in vitro and in vivo.
Finally, we aimed to develop a chemically defined ECM in which to culture hPOs as the commonly used ECMs (Matrigel and BME 2-RGF) are animal-derived and are unsuitable for cell therapies. In collaboration with Cellendes, a company specialised in hydrogel technology, we could show for the first time the derivation and expansion of hPOs in a fully chemically defined hydrogel.
Our demonstration of the long-term expansion of human pancreas organoids in chemically defined conditions makes this an exciting system for many areas of research (Figure 3). This system can be used as a model of healthy ductal pancreas biology and the development of disease models of the ductal compartment, such as cystic fibrosis. Organoids can also be derived from diseased states and have already been used to model different types of pancreatic cancer (PDAC and IPMN).
[caption id="attachment_11731" align="aligncenter" width="605"]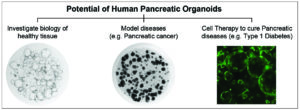 Figure 3: Human pancreatic organoids have the potential to be used in many areas of research such as investigations into healthy tissue, diseased tissue and regenerative medicine.[/caption]
Critically, our protocol does not rely on altering the genome or identity of the cells in order to achieve long-term expansion. This, combined with the lack of tumour formation when transplanted into mice, makes hPOs a safe approach for potential cell therapies. Cell therapies offer hope for patients without available curative treatment options. They provide the possibility of treating a disease by extracting cells from the patient or other donors, altering them in the lab to generate working cells and transplanting them into the patient. In theory, this would treat the disease while removing the need for immunosuppression as the cells would be either self-derived or very closely matched to the patient.
The adaptation of the culture system for human cells was funded by an EU programme (LSFM4LIFE) which aims at investigating the potential of hPOs as a clinical cell therapy. Whether hPOs could be the source for a cell therapy transplantation for diabetes remains to be investigated.
Figure 3: Human pancreatic organoids have the potential to be used in many areas of research such as investigations into healthy tissue, diseased tissue and regenerative medicine.[/caption]
Critically, our protocol does not rely on altering the genome or identity of the cells in order to achieve long-term expansion. This, combined with the lack of tumour formation when transplanted into mice, makes hPOs a safe approach for potential cell therapies. Cell therapies offer hope for patients without available curative treatment options. They provide the possibility of treating a disease by extracting cells from the patient or other donors, altering them in the lab to generate working cells and transplanting them into the patient. In theory, this would treat the disease while removing the need for immunosuppression as the cells would be either self-derived or very closely matched to the patient.
The adaptation of the culture system for human cells was funded by an EU programme (LSFM4LIFE) which aims at investigating the potential of hPOs as a clinical cell therapy. Whether hPOs could be the source for a cell therapy transplantation for diabetes remains to be investigated.
The slow loris (Nycticebus spp.) is the world’s only venomous primate, just one of seven venomous species of mammal.
From a defensive posture, with raised arms, it can produce a potent venom by combining fluid from its brachial gland with saliva. The toxic mixture is either applied to the top of the head to ward off attackers or kept in the mouth to administer with a sharp and painful bite. Slow loris venom has been known to cause anaphylaxis and even death in small animals and humans.
Although folklore in Southeast Asia has long ascribed deadly or even black magic properties to the slow loris and its blood or venom, the first scientific reports of Nycticebus venom only emerged in the mid-1990s.
Chemical structure of slow loris venom
The authors review a series of experiments to shed light on the biochemistry of slow loris venom. High performance liquid chromatography was used to identify the various components of the loris’ brachial gland secretions. The technique identified a protein with strong genetic similarity to the cat allergen Fe1-d1.
Fe1-d1 is used by cats as a species recognition molecule. If the brachial secretions in slow loris venom can be used by the animals in this way, it seems likely that the venom may have utility beyond that of a direct toxic defense.
Function of slow loris venom
There is a related hypothesis that slow loris venom was initially used for species recognition – not unlike Fe1-d1 in cats – to stake territorial claims or to act as a warning signal to other lorises, and eventually evolved into a potent direct venom.
Observations in the lab and in field studies suggest that the slow loris uses its venom to directly repel predators, especially those reliant on smell (e.g. cats and sun bears). Anointment of its fur with venom also provides the loris with a first line chemical defense against parasites.
Nekaris and colleagues also review evidence for venom being used by slow lorises against their conspecifics. Intersexual competition and territorial skirmishes occur frequently between lorises, especially in breeding season, and these fights are often brought to a close by venom-laden bites. In this context, anointment of fur on the loris’ head with venom is a measure to protect the most vulnerable part of their body during fights.
Why did the venom evolve?

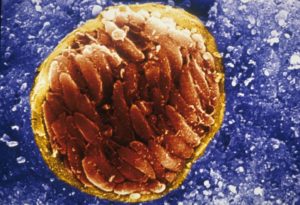 A scanning electron micrograph of Toxoplasma gondii, tissue cyst in brain of an infected mouse: Credit: David Ferguson[/caption]
That surrounding bradyzoites, is less permeable than the wall enclosing tachyzoites and is a heavily glycosylated structure with many stage-specific proteins of unknown function inserted in it. In addition, unlike tachyzoites, bradyzoites undergo anaerobic respiration and their metabolism is slower. Bradyzoite formation leads to a latent infection that lasts a lifetime and can result in recrudescence that can be life threatening in immunocompromised individuals.
The presence of this dormant stage greatly enhances transmission of the parasite (if tissues containing bradyzoites are eaten) thus making an understanding of the formation of this stage important.
Differentiation of tachyzoites into bradyzoites can be induced in vitro by a variety of stressors, and differences in the regulation of a large number of genes have been recognised. However, the differentiation process is poorly understood.
Differentiation investigated
Members of the Sebastian Lourido lab at the Whitehead Institute and MIT have developed a pooled genetic screen using CRISPR mediated disruption of Toxoplasma genes. This has enabled them to identify and investigate fitness conferring genes. Their technique is elegantly explained by Sebastian Lourido in two videos available from his profile.
In a recently published paper his group report that they used this protocol to look for mutant Toxoplasma, deficient in differentiation. To do this they produced a reporter strain that expresses a fluorescent marker when bradyzoites were formed; Cas9 allowed efficient gene inactivation.
After stressing their cell cultures, and using a cell sorter to separate bradyzoites from tachyzoites, they found 1,311 genes were upregulated and 933 genes were downregulated in bradyzoites. Two gene libraries were constructed and screened for genes required for bradyzoite formation.
A master regulator
The caspase-mediated screening allowed them to identify a single gene as essential for differentiation. They named it Bradyzoite-Formation Deficient 1 (BFD1) and found it to be conserved among cyst-forming protozoan parasites in the subclass Coccidia. Knockout of this gene resulted in tachyzoites that grew normally in culture, but did not differentiate into bradyzoites when cultures were stressed. When mice were infected with ΔBFD1 parasites, no bradyzoite cysts were formed in the brain.
The authors found that BDF1 binds near the transcriptional start sites of many genes upregulated during differentiation, and conditional expression induced differentiation without the need to stress tachyzoite cultures. The authors conclude that BDF1 is a master regulator for chronic-stage differentiation in Toxoplasma.
This paper, published in January in Cell, is a model exposition of the methodology used and the thinking behind the discovery of this key to understanding how a transmission stage of such a common parasite is formed. The authors point out the biotechnological potential for ΔBFD1, and for ΔBDT1 parasites as an attenuated vaccine strain. Let us hope this bears fruit.
A scanning electron micrograph of Toxoplasma gondii, tissue cyst in brain of an infected mouse: Credit: David Ferguson[/caption]
That surrounding bradyzoites, is less permeable than the wall enclosing tachyzoites and is a heavily glycosylated structure with many stage-specific proteins of unknown function inserted in it. In addition, unlike tachyzoites, bradyzoites undergo anaerobic respiration and their metabolism is slower. Bradyzoite formation leads to a latent infection that lasts a lifetime and can result in recrudescence that can be life threatening in immunocompromised individuals.
The presence of this dormant stage greatly enhances transmission of the parasite (if tissues containing bradyzoites are eaten) thus making an understanding of the formation of this stage important.
Differentiation of tachyzoites into bradyzoites can be induced in vitro by a variety of stressors, and differences in the regulation of a large number of genes have been recognised. However, the differentiation process is poorly understood.
Differentiation investigated
Members of the Sebastian Lourido lab at the Whitehead Institute and MIT have developed a pooled genetic screen using CRISPR mediated disruption of Toxoplasma genes. This has enabled them to identify and investigate fitness conferring genes. Their technique is elegantly explained by Sebastian Lourido in two videos available from his profile.
In a recently published paper his group report that they used this protocol to look for mutant Toxoplasma, deficient in differentiation. To do this they produced a reporter strain that expresses a fluorescent marker when bradyzoites were formed; Cas9 allowed efficient gene inactivation.
After stressing their cell cultures, and using a cell sorter to separate bradyzoites from tachyzoites, they found 1,311 genes were upregulated and 933 genes were downregulated in bradyzoites. Two gene libraries were constructed and screened for genes required for bradyzoite formation.
A master regulator
The caspase-mediated screening allowed them to identify a single gene as essential for differentiation. They named it Bradyzoite-Formation Deficient 1 (BFD1) and found it to be conserved among cyst-forming protozoan parasites in the subclass Coccidia. Knockout of this gene resulted in tachyzoites that grew normally in culture, but did not differentiate into bradyzoites when cultures were stressed. When mice were infected with ΔBFD1 parasites, no bradyzoite cysts were formed in the brain.
The authors found that BDF1 binds near the transcriptional start sites of many genes upregulated during differentiation, and conditional expression induced differentiation without the need to stress tachyzoite cultures. The authors conclude that BDF1 is a master regulator for chronic-stage differentiation in Toxoplasma.
This paper, published in January in Cell, is a model exposition of the methodology used and the thinking behind the discovery of this key to understanding how a transmission stage of such a common parasite is formed. The authors point out the biotechnological potential for ΔBFD1, and for ΔBDT1 parasites as an attenuated vaccine strain. Let us hope this bears fruit.
One particularly compelling hypothesis for slow loris venom etiology is explored in depth by the authors: the theory that slow loris venom may have evolved to mimic cobras.
The facial markings of various slow loris species bear a strong resemblance to the dorsal patterns of the spectacled cobra (Naja naja). Other serpentine features have been observed in the slow loris, from its hiss-like vocalizations when threatened to the graceful movement conferred to the loris by its additional vertebrae.
Nekaris and colleagues suggest that these features could reflect a form of Müllerian mimicry in the slow loris. The spectacled cobra would be unwanted and noxious prey for a number of predators. For the predator looking to identify its prey, the slow loris’ resemblance to these cobras would bring uncertainty and risk to any attack.
Savannah-like conditions brought on by climatic events in Southeast Asia several million years ago would have exposed the slow loris to a number of new predators in common with the spectacled cobra. This may have triggered a selection event in the species, towards cobra mimicry.
Endangered status
The Javan slow loris (Nycticebus javanicus) has been listed as endangered, with a suspected decline of at least 50% over the last three generations. Slow lorises have been the target of capture for a burgeoning illegal pet trade, fuelled in part by their misrepresentation in viral video content.
Thankfully, awareness of the slow loris’ endangered status and illegal trade seems to be increasing – even amid the quagmire of Youtube comments – as the result of educational efforts by conservation scientists and outreach groups.
About the authors
- Anna Nekaris is Professor in Anthropology and Primate Conservation at Oxford Brookes University, and the director of the Little Fireface Project, named after the Sudanese word for the slow loris. She is also active on Twitter and has featured in numerous interviews and discussion panels on the slow loris.
- Dr Richard Moore is research director of International Animal Rescue’s slow loris rescue centre in Indonesia.
- Johanna Rode conducted her PhD research on the ecological function of slow loris venom in Java.
- Dr Brian Fry is group leader of the Venomics Laboratory at the School of Biological Sciences, University of Queensland, and a world expert in all manner of venomous animals (from spiders and snakes to jellyfish and sharks).
Article information
Mad, bad and dangerous to know: the biochemistry, ecology and evolution of slow loris venom
Anne-Isola Nekaris, Richard S Moore, Johanna Rode and Bryan G Fry
Journal of Venomous Animals and Toxins including Tropical Diseases 2013, 19:21
doi:10.1186/1678-9199-19-21
full text

Comments